Suomalaisia prekliinisiä liikehäiriötutkimuksia 2024
Vuonna 2024 ilmestyi parikymmentä liikehäiriöalan kokeellista tutkimusta, joissa suomalaisia oli mukana. Määrä on ollut samaa luokkaa viime vuosina. Merkittävä osa käsitteli hermokasvutekijöitä tai ⍺-synukleiinia (a-Syn) tavalla tai toisella, ja katsausartikkeleita oli monia. Kapea tutkimuskirjo on huolestuttavaa, samoin ehkä kansainvälisten tutkijoiden suuri määrä. Missä luuraavat suomalaiset perustutkijat? Etenkin lääkäritutkijat ovat hupeneva luonnonvara.
Ihmisen kantasoluista erikoistuneisiin soluihin
Lehtosen ryhmä Kuopiosta (AIV) on edelleen ollut hyvin aktiivinen, mutta kaikissa 3 raportissa perusasetelma on sama, vain kohdesolu on kaikissa eri. Tutkimusryhmä on tuottanut kantasoluja sekä LRKK2-mutaatioita kantavista Parkinson-(PD)-potilaista että terveistä kontrolleista ja sitten erilaistanut nämä mikrogliaksi, endoteelisoluiksi tai perisyyteiksi. Solutyypit on varmistettu monipuolisesti. Nykyisin kantasolujan saa myös ostaa kaupallisesti. Perusajatuksena on se, että PD:ssa häiriintyvät tai vaurioituvat myös monet muut solutyypit kuin dopamiinisolut.
Niskanen ym (17) tuotti aktiivisia mikrogliasoluja. Niiden asianmukaisuus tarkistettiin moninaisilla kokeilla ja oikeiksi havaittiin. Mikroglian toiminnassa oli suuria eroja PD-potilaiden ja terveiden välillä. PD-potilaiden mikroglia otti huonosti sisäänsä ulkoisesti annettua a-Syn fibrillivalmistetta eli niiden fagosytoosi oli heikentynyt. Tulehdus vielä huononsi tilannetta, lisäsi sytokiinien ja typpioksidin tuotantoa. Myös mitokondrioien toiminta huononi. Ovatko huonosti voivat mikroglia syy vain seuraus?
Sonninen ym. (23) tuotti endoteelisoluja Lehtosen ryhmän menetelmillä potilaista ja terveistä. Endoteelisolut ovat keskeisiä mm. veri-aivo-esteen (BBB) muodostamisessa ja toiminnassa. A-Syn-kertymät olivat merkittävän suuria PD-potilaiden endoteelisoluissa, joiden metabolinen aktiivisuus heikkeni vähän (tilastollisesti kyllä). Muutenkin soluissa havaittiin PD:lle tyypillisiä biokemiallisia muutoksia. Keinotekoinen tulehdus (tumor necrosis factoria annettiin) pahensi tilannetta, etenkin tulehduksen pidentyessä. BBB:n vuotamista ei kuitenkaan havaittu.
Peltonen ym. (19) tuotti ym. tekniikoilla kantasoluista perisyyttejä. Nämä vaikeasti määriteltävät solut säätelevät mm. verisuonia ja tulehdusreaktioita aivoissa ja ovat muodostamassa BBB:iä endoteelisolujen kanssa. Yhteistoimintaa näillä soluilla on muutenkin eikä niitä ole aina helppo edes erottaa toisistaan. Perisyytit eivät kerää a-Syn-säikeitä sisäänsä, mutta PD-potilaiden perisyytit liikkuvat huonosti ja erittävät tulehdusta välittäviä aineita, etenkin stimuloitaessa. Yhteys PD:iin jää hyvin ohueksi.
Lehtosen ryhmän Albert ym. (2) tarkastelee lyhyessä katsauksessa gliasolusiirtojen hoidollista potentiaalia aivojen rappeumasairauksien hoidossa. Yhtenä kapeana indikaationa on PD, jossa ns. MitoPark-hiirimallia on käytetty asian tutkimiseen. Pääpaino on muissa aivosairauksissa. Siirtoja on tehty sekoituksella, jossa on makrofageja ja gliasoluja, ja on annosteltu itse asiassa GDNF-kasvutekijää. Kaikki vaikuttaa olevan ”wishful thinking”-vaiheessa.
Ihmisen kantasoluja on käytetty myös Kapucun ym. tutkimuksessa (11; ks. myöhemmin).
Hermokasvutekijätutkimukset
Voutilaisen ryhmä jatkaa perusteellisia ja suuritöisiä tutkimuksiaan. Nam ym. (17) suorittivat ihmisen CDNF-hermokasvutekijän siirtoja AAV-viruksen avulla MPTP-hiirimallissa. Tulokset ovat selvät ja vahvistavat mm. allekirjoittaneen aiempia tuloksia rotan 6-OHDA-mallissa (Bäck ym. 2013:Brain Behav. 3,75–88. https:// doi. org/ 10. 1002/ brb3. 117). Infuusio annettiin keskelle striatumia ja hCDNF ilmeni sekä striatumissa että S. nigrassa, hiirien suoriutuminen ja tasapaino paranivat merkitsevästi eikä toksisuutta ilmaantunut. Aivan kontrollihiirien tasolle ei kuitenkaan päästy. hCDNF levisi hiukan pienemmälle alueelle kuin pelkkä väriaine. Tulehdusvälittäjien määrä väheni. Keskeisen GRP78-säätelyvalkuaisen määrä astrogliassa alentui.
Andressoon ryhmä on myös ollut aktiivinen ja hänen lähestymistapansa hyvin omaperäinen. Montaño-Rodriquez ym. (16) selvittivät varsin keskeistä ongelmaa: miksi GDNF on niin tehokas neuroprotektori, vaikka se ilmenee vain 0.6%:ssa striatumin dopamiinisoluista (lyhyissä interneuroneissa)? Sopivien vasta-aineiden puuttuessa he käyttivät knock-in-tekniikkaa RET-reseptorin visualisointiin. Havaittiin, että GDNF:ää sisältävät hermot ”imevät” RET-positiivisia neuroneita jopa 7 kertaa kauempaa kuin muita (ns. medium spiny neurons), joita on jopa 97% kaikista. Tämä pieni GDNF-populaatio vastaa suureksi osaksi dopamiinituotannosta ja myös dopamiinihermojen haaromisesta. Havainto selittää ensi kertaa, miksi varsin pieneltä vaikuttava GDNF-efekti on toiminnallisesti niin tehokas. Erittäin fiksu ja tarpeellinen työ.
Saarman ryhmällä on jälleen lukuisia raportteja hermokasvutekijöistä, mutta yhtä lukuun ottamatta (6) kaikki ovat katsauksia.
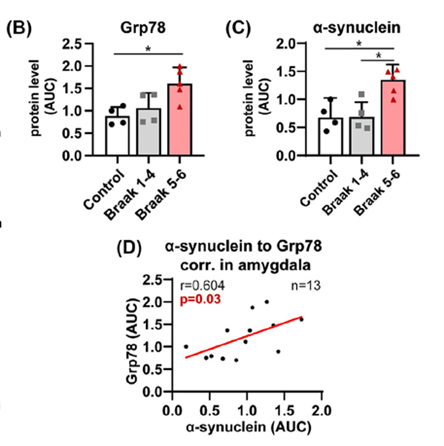
Hrabos ym. (6) tutkivat Oxfordin aivopankista 45 vainajan aivot, joista 12 oli ollut aivoiltaan terveitä ja 33:lla oli ollut pysyvä tai ajoittain oireileva Lewy-kappaletauti. Monilta aivoalueilta mitattiin mm. hermokasvutekijöitä (MANF, CDNF), liukoista a-Syn:iä ja laskostamattomien proteiinien reaktioon (UPR) liittyviä säätelytekijöitä (GRP78 ja eIF2). Tuloksena oli, että UPR-merkkiaineet lisääntyivät liukoisen a-Syn:n nousun myötä useilla aivoalueilla, mm. amygdalassa ja dopamiinihermoissa, mutta nousu ei riippunut lainkaan Lewy-patologiasta. Kuvassa korrelaatio on esitetty amygdalan alueelta. MANF:in tai CDNF:n pitoisuuksissa ei ollut eroja eri ryhmien välillä. MANF:n pitoisuus pieneni ikääntyessä. Tämä oikeilla ihmisaivoilla tehty peripohjainen analyysi osoitti, etteivät Lewy-kappaleet suoraan liittyneet lainkaan hermoston rappeutumiseen.
Saarman ryhmän katsaukset ovat kaikki rappeumasairauksien hoitoon liittyviä. Airavaara ja Saarma (1) pohtivat käsikirjatasolla aivosairauksien kasvutekijähoitoja annostelutavan perusteella. Virustransfektiot vaiko suorat infuusiot? Barkerin ym. (3) aiheena on äskeisen kasvitekijäkokouksen anti, josta ei ole vielä tekstiä saatavilla. Kakinen ym. (9) keskustelevat laajalla skaalalla nanolääketieteen keinoja aivosairauksien hoidossa, ja PD on vain pienessä määrin mukana. Teorioita on lukuisia, käytännön sovelluksia tuskin lainkaan. BBB on edelleen erittäin kova vastus kaikille keskushermostosairauksien hoidoille.
Lohelaid ym. (15) katsaus tarkastelee jälleen kerran CDNF:n vaikutusmekanismeja. CDNF eroaa muista hermokasvutekijöistä siinä, että se sisältää ns. KDEL-signaalin, joka ohjaa sen solunsisäiseen verkostoon (ER, endoplasmic reticulum). Näin ollen ER-stressin hillintä on yksi ilmeinen vaikutustapa. Se ei sulje pois muita vaikutusmekanismeja kuten a-Syn-kertymisen estäminen. ER-stressiin liittyy olennaisesti säätelevä GRP78-proteiini, jonka stressi irrottaa ER-sensorista, jolloin UPR-signaali laukeaa (laskostumattomien proteiinin reaktio). Toimiiko tämä suoraan juuri näin PD:n synnyssä, on vielä todistamatta.
Huttusen ryhmä tai Herantis ei raportoi mitään uutta kliinisiin kokeisiin menneestä CDNF-johdoksesta HER-096. Sen sijaan Graewert ym. (5) tutkivat laajassa kansainvälisessä tutkimuksessa CDNF:n ja ym. GRB78-proteiinin interaktioita fysikokemiallisin menetelmin. GRP78 todistettiin keskeiseksi säätelytekijäksi laskostumattomien proteiinin haitallisessa reaktiossa. Tähän ovat monet tutkijat päätyneet muita teitä (esim. 6 ja 15 tässä katsauksessa). Tulos varmistettiin myös pistemutaatioilla, jolloin ilmeni, että yksikin mutaatio GBR78:n rakenteessa poisti CDNF:n suojaavan vaikutuksen.
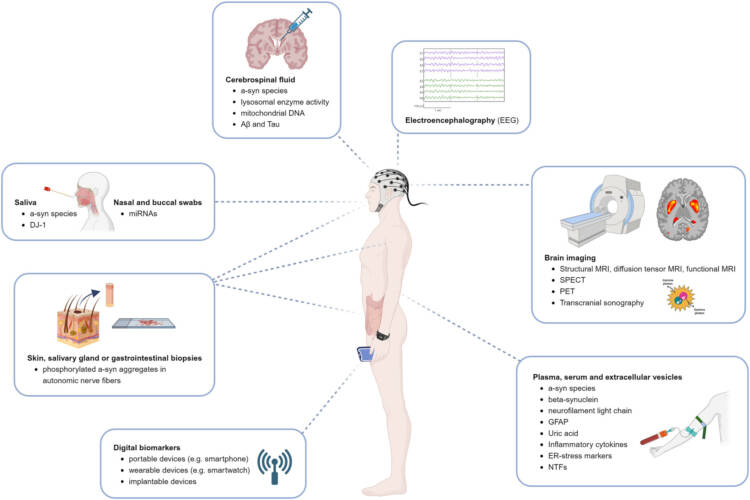
Tuominen ja Renko (24) tekevät kattavan yhteenvedon biomarkkereista, joita voidaan käyttää PD:n varhaisen vaiheen diagnostiikassa, erityisesti ajatellen mahdollisia hermokasvu-tekijähoitoja. Niiden kliininen teho on ollut vaisu ehkä johtuen jo pitkälle edenneestä taudista. Hoitoja pitäisi antaa jo ennen varsinaisia vakavia oireita. Tämä on vaikea yhtälö, jos kasvutekijöiden anto edellyttää aivoleikkausta. Tilanne on aivan toinen, jos aivojen ulkopuolinen hoito onnistuu (vrt. Herantiksen HER-096). Monenlaista varhaista diagnostiikkaa voidaan tehdä biokemiasta aina kuvantamiseen saakka. Yksinkertaisen verikokeen mahdollisuus näyttää lisääntyneen (esim. kevyiden neurofibrillien määrä). Julkaisun kuva kuvaa monipuolisesti eri mahdollisuuksia, joita neurologeilla on.
MIRAS
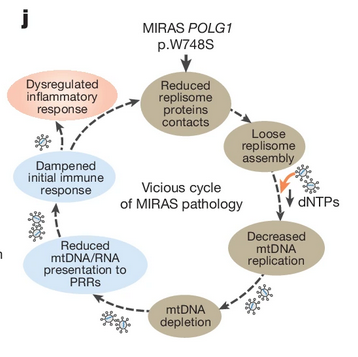
Uutta ja yllättävää tietoa MIRAS ataksiasta julkaistiin Nature-tiedelehdessä oudossa yhteydessä: tutkittaessa viruslääkeresistenssiä. MIRAS (mitokondriaalinen peittyvä ataksia) johtuu POLG-entsyymin mutaatiosta p.W7485. POLG:n (DNA polymeraasi gamma 1) aktiviteettiä säätelevät tietyt virusinfektiot ja ym. mutaatio herkistää säätelyä. Hyvin perusteellisessa ja monimutkaisessa tutkimussarjassa Kang ym. (10) Anu Suomalaisen ryhmästä tekivät MIRAS-hiirimallin knock-in tekniikalla, siis siirtämällä p.W7485-mutaation hiiriin. Tietty virusinfektio aiheutti monivaiheisen biologisen häiriön kautta mitokondriaalisen RNA:n toiminnan heikkoutta, heikensi immuniteettiä ja voimisti infektoivan viruksen vaikutusta, jolloin MIRAS-oireet ilmaantuivat: GABA-neuronien tuhoa, kouristuksia, maksavaurioita. Synnynnäiseen immuunimekanismiin puuttuminen saattaisi tarjota hoidollisen kohteen mitokondriosairauksille.
Sekalaisia tutkimuksia
Turconi ym. (25) puolestaan tutkivat nikotiiniribosidin (käytännössä B3-vitamiinin) tehoa kahdessa PD-mallissa. C. elegans madossa, joka yliekspressoi a-Syn:iä ja hiirellä laktokystiinimallissa (ns. proteosomimekanismi eli mitokondriovauriomalli). Tulokset hämmentävät. Matomallissa B3 toimi suojaavasti, mutta hiirimallissa kohtuullinen suojavaikutus vaihtuikin dopamiinisynteesin laskuksi parin viikon kuluessa. Mitokondriomuutokset olivat melko lailla odotettuja, mutta eivät siis korreloineet hiirien käyttäytymiseen. Vitamiinihoito ei taida sekään olla ratkaisu PD:n hoitoon. Tampereen yliopistolla on kehitetty kolmiosastoinen in vitro-laitteisto, jossa voidaan viljellä ihmisen kantasoluista tuotettuja dopamiinisoluja (Kapucu ym. 11). Laitteistoon on kytketty mikrochippejä ja mikroelektrodeja erilaisia mittauksia varten. Osastot ovat yhteydessä toisiinsa mikrotunneleillla, joiden läpi esim. aksonit voivat kasvaa. Laitteistolla on tutkittu mm. a-Syn:n sakkautumista ja leviämistä käyttäen tunnettua fibrillimallia. Ca++-virtoja, mitokondrioiden liikkeitä ja hermosolujen aktiviteettiä on mitattu on-line. Jopa parin viikon seurannassa neuronit voivat hyvin, mutta mitokondrioiden toiminta häiriintyi ja Ca++-tasot alenivat. Glutamaattijärjestelmään oli vain lievä vaikutus. Kaikkiaan muutokset olivat vähäisiä, vaikka fibrillialtistus sinänsä oli hyvin nähtävissä. Laitteisto soveltuu jatkotutkimuksiin, joista ei todellakaan ole puutetta.
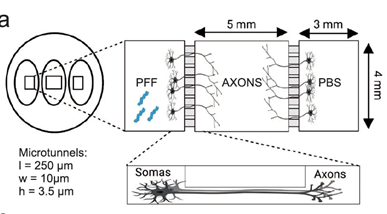
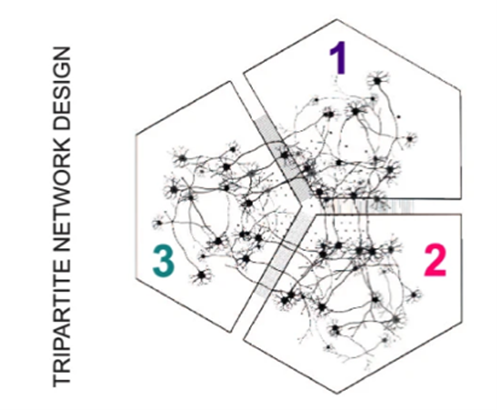 Kuva 4. Engineered circular tripartite and axon elongation cortical models. Julkaisu 11, kuva 2A-D
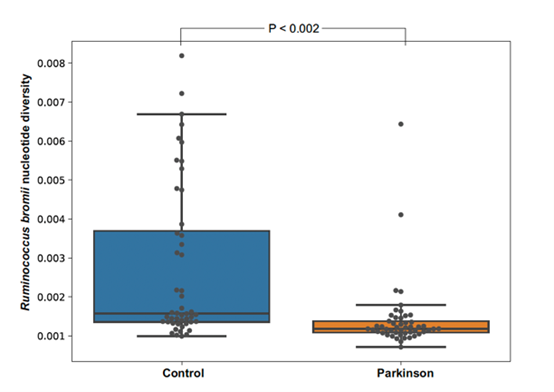
Kuva 4. Engineered circular tripartite and axon elongation cortical models. Julkaisu 11, kuva 2A-DDuru ym. (4) tekivät mahdottomalta tuntuvan työn suoliston bakteerien parissa selvittäen ns. metagenomi-assembled genomit (MAGs) terveillä (n=68) ja PD-potilailla (n=68). Peräti 952 MAG:tä rakennettiin. Lukemattomista bakteerinimistä mainittakoon vain Ruminococcus bromii, mutta lopputulema oli se, että terveiden henkilöiden suolessa bakteerisukujen diversiteetti oli laajempi kuin PD-potilaiden suolessa.
Puolalais-Kuopiolainen katsaus (Jastrzębski ym. 8) tarkastelee hajanaista ryhmää ”pienmolekyyylejä”, joilla saattaisi olla merkitystä neurogeneesissä tai ehkä neuroplastisuudessa. Lähtökohtana pidetään aivosairauksien perusmekanismeja ja toisaalta sitä, että osassa aivoja on muokattavissa olevia hermosoluja. Kovin syvälle ei päästä. Luettelonomaisesti esitellään kirjava joukko yhdisteitä, esim. fluoksetiini, litium, formoteroli, pregnenololi, harmiini ja joukko koodinimiä. Kirjoittajat luettelevat myös joukon erilaisia biologisten prosessien välittäjiä, ”third messengers”, mutta eri yhdisteiden yhteys näihin jää ohueksi. Julkaisu on Acta Pharmacologica Sinica, mikä selittänee jotakin.
Koistinahon katsauksia ilmestyy vuosittain ja lisäksi hän on edelleen mukana useissa Lehtosen julkaisuissa ainakin solujen lahjoittajana. Koistinahon omissa katsauksissa Kettunen ym. (12) käsittelee sekä aivojen että periferian tulehdusten merkitystä rappeumasairauksissa. PD:n kohdalla painotetaan yhteyttä useisiin immunovastetta välittäviin tekijöihin ja mutaatioihin, joita tulehdukset provosoivat. Yleinen johtopäätös on, että tulehduksilla on merkitystä, mutta tulehduksen laadulla tai sijainnilla ei niinkään. LRRK2-mutaation yhteys esim. reoviruksen kuolinriskiä lisääjänä tuodaan esille yhtenä esimerkkinä. Myös aivojen ulkopuoliset tulehdukset (Helicobacter pylori, suoliston muut tulehdukset, ientulehdukset) voivat toki liittyä jotenkin aivosairauksiin. Muutoin tämän katsauksen anti on vähäinen ilman syvällisiä oivalluksia.
Lähinnä anekdotaalinen on myös Keutersin ym. (13) raportti, että PD-lääke benseratsidi suurina pitoisuuksina voisi olla hyödyllinen aivohalvauksissa, tuskin PD:ssa.
KM Huttunen (7) on pyydetyssä katsauksessa (”expert opinion”) lausunut mielipiteitään prodrug-teknologiasta ajatellen lääkeaineiden aivoihin saattamista. PD:n lääkkeistä ei ole kuin pari esimerkkiä, mm. dopamiinin ja glukoosin muodostama hydridi, ei edes levodopaa ole käsitelty.
Saloranta ym. (21) tutkivat kahden PD-lääkkeen (ropiniroli, apomorfiini) koirien oksetusta aiheuttamaan ilmeisesti myrkytysten hoitoa ajatellen. Mitään muuta PD:iin liittyvää tässä ei ole.
Kirjoittaja on Pekka T. Männistö, Farmakologian ja lääkekehityksen emeritusprofessori, Suomen Parkinson-säätiön hallituksen asiantuntijajäsen
Kirjallisuus
(* Rahoitusta Suomen Parkinson-säätiöltä)
1: Airavaara M, Saarma M. Viral and nonviral approaches. Handb Clin Neurol.
2024;205:83-97. doi: 10.1016/B978-0-323-90120-8.00008-3. PMID: 39341664.
2: Albert K, Goldsteins G, Kälvälä S, Jolkkonen J, Lehtonen Š. Glial cell transplant for brain diseases: the supportive saviours? Translational Medicine Communications. 2024 Jul 13;9(1):22.
3: Barker RA, Saarma M, Svendsen CN, Morgan C, Whone A, Fiandaca MS, Luz M, Bankiewicz KS, Fiske B, Isaacs L, Roach A. Neurotrophic factors for Parkinson’s disease: Current status, progress, and remaining questions. Conclusions from a 2023 workshop. Journal of Parkinson’s Disease. 2024:1877718X241301041.
4: Duru IC, Lecomte A, Shishido TK, Laine P, Suppula J, Paulin L, Scheperjans F, Pereira PA, Auvinen P. Metagenome-assembled microbial genomes from Parkinson’s disease fecal samples. Scientific Reports. 2024 Aug 14;14(1):18906.
5: Graewert MA, Volkova M, Jonasson K, Määttä JAE, Gräwert T, Mamidi S,
Kulesskaya N, Evenäs J, Johnsson RE, Svergun D, Bhattacharjee A, Huttunen HJ.
Structural basis of CDNF interaction with the UPR regulator GRP78. Nat Commun.
2024 Sep 18;15(1):8175. doi: 10.1038/s41467-024-52478-0. PMID: 39289391; PMCID:
PMC11408689.
6: Hrabos D, Poggiolini I, Civitelli L, Galli E, Esapa C, Saarma M, Lindholm P,
Parkkinen L. Unfolded protein response markers Grp78 and eIF2alpha are
upregulated with increasing alpha-synuclein levels in Lewy body disease.
Neuropathol Appl Neurobiol. 2024 Aug;50(4):e12999. doi: 10.1111/nan.12999. PMID:
39036837.
7: Huttunen KM. Improving drug delivery to the brain: the prodrug approach.
Expert Opin Drug Deliv. 2024 May;21(5):683-693. doi: 10.1080/17425247.2024.2355180. Epub 2024 May 16. PMID: 38738934.
8: Jastrzębski MK, Wójcik P, Stępnicki P, Kaczor AA. Effects of small molecules
on neurogenesis: Neuronal proliferation and differentiation. Acta Pharm Sin B.
2024 Jan;14(1):20-37. doi: 10.1016/j.apsb.2023.10.007. Epub 2023 Oct 20. PMID:
38239239; PMCID: PMC10793103.
9: Kakinen A, Jiang Y, Davis TP, Teesalu T, Saarma M. Brain Targeting
Nanomedicines: Pitfalls and Promise. Int J Nanomedicine. 2024 May
27;19:4857-4875. doi: 10.2147/IJN.S454553. PMID: 38828195; PMCID: PMC11143448.
10: Kang Y, Hepojoki J, Maldonado RS, Mito T, Terzioglu M, Manninen T, Kant R,
Singh S, Othman A, Verma R, Uusimaa J, Wartiovaara K, Kareinen L, Zamboni N,
Nyman TA, Paetau A, Kipar A, Vapalahti O, Suomalainen A. Ancestral allele of DNA
polymerase gamma modifies antiviral tolerance. Nature. 2024
Apr;628(8009):844-853. doi: 10.1038/s41586-024-07260-z. Epub 2024 Apr 3. PMID:
38570685; PMCID: PMC11041766.
11: Kapucu FE, Tujula I, Kulta O, Sukki L, Ryynänen T, Gram H, Vuolanto V,
Vinogradov A, Kreutzer J, Jensen PH, Kallio P, Narkilahti S. Human tripartite
cortical network model for temporal assessment of alpha-synuclein aggregation
and propagation in Parkinson’s Disease. NPJ Parkinsons Dis. 2024 Jul
28;10(1):138. doi: 10.1038/s41531-024-00750-x. PMID: 39069518; PMCID:
PMC11284226.
12: Keuters MH, Keksa-Goldsteine V, Rõlova T, Jaronen M, Kettunen P, Halkoluoto
A, Goldsteins G, Koistinaho J, Dhungana H. Benserazide is neuroprotective and
improves functional recovery after experimental ischemic stroke by altering the
immune response. Sci Rep. 2024 Aug 2;14(1):17949. doi:
10.1038/s41598-024-68986-4. PMID: 39095453; PMCID: PMC11297251.
13: Kettunen P, Koistinaho J, Rolova T. Contribution of CNS and extra-CNS infections to neurodegeneration: a narrative review. Journal of Neuroinflammation. 2024;21.
14: Ei käytössä
15: Lõhelaid H, Saarma M, Airavaara M. CDNF and ER stress: Pharmacology and
therapeutic possibilities. Pharmacol Ther. 2024 Feb;254:108594. doi:
10.1016/j.pharmthera.2024.108594. Epub 2024 Jan 28. PMID: 38290651.
16: Montaño-Rodriguez AR, Schorling T, Andressoo JO. Striatal GDNF Neurons
Chemoattract RET-Positive Dopamine Axons at Seven Times Farther Distance Than
Medium Spiny Neurons. Cells. 2024 Jun 19;13(12):1059. doi:
10.3390/cells13121059. PMID: 38920687; PMCID: PMC11202212.
17: Nam J, Richie CT, Harvey BK, Voutilainen MH. Delivery of CDNF by AAV-
mediated gene transfer protects dopamine neurons and regulates ER stress and
inflammation in an acute MPTP mouse model of Parkinson’s disease. Sci Rep. 2024
Jul 17;14(1):16487. doi: 10.1038/s41598-024-65735-5. PMID: 39019902; PMCID:
PMC11254911
18: Niskanen J, Peltonen S, Ohtonen S, Fazaludeen MF, Luk KC, Giudice L,
Koistinaho J, Malm T, Goldsteins G, Albert K, Lehtonen Š. Uptake of alpha-
synuclein preformed fibrils is suppressed by inflammation and induces an
aberrant phenotype in human microglia. Glia. 2024 Oct 22. doi:
10.1002/glia.24626. Epub ahead of print. PMID: 39435593.
19: Peltonen S, Sonninen TM, Niskanen J, Koistinaho J, Ruponen M, Lehtonen Š.
Mutated LRRK2 induces a reactive phenotype and alters migration in human iPSC-
derived pericyte-like cells. Fluids Barriers CNS. 2024 Nov 18;21(1):92. doi:10.1186/s12987-024-00592-y. PMID: 39551752; PMCID: PMC11571670.
20: Ei käytössä
21: Saloranta LI, Levijoki JM, Vuorela AM. An experimental study of consecutive administration of ropinirole and apomorphine for emesis induction in dogs. Journal of Veterinary Emergency and Critical Care. 2024 Jan;34(1):31-9.
*22: Singh A, Panhelainen A, Reunanen S, Luk KC, Voutilainen MH. Combining
fibril-induced alpha-synuclein aggregation and 6-hydroxydopamine in a mouse
model of Parkinson’s disease and the effect of cerebral dopamine neurotrophic
factor on the induced neurodegeneration. Eur J Neurosci. 2024 Jan;59(1):132-153.
doi: 10.1111/ejn.16196. Epub 2023 Dec 10. PMID: 38072889.
23: Sonninen TM, Peltonen S, Niskanen J, Hämäläinen RH, Koistinaho J, Lehtonen Š. LRRK2 G2019S Mutated iPSC-Derived Endothelial Cells Exhibit Increased α-Synuclein, Mitochondrial Impairment, and Altered Inflammatory Responses. International Journal of Molecular Sciences. 2024 Nov 29;25(23):12874.
24: Tuominen RK, Renko JM. Biomarkers of Parkinson’s disease in perspective of
early diagnosis and translation of neurotrophic therapies. Basic Clin Pharmacol
Toxicol. 2024 Sep;135(3):271-284. doi: 10.1111/bcpt.14042. Epub 2024 Jul 8.
PMID: 38973499.
*25: Turconi G, Alam F, SenGupta T, Pirnes-Karhu S, Olfat S, Schmidt MS, Mätlik
K, Montaño-Rodriguez A, Heiskanen V, Garton D, Piepponen PT, Brenner C, Holmberg
CI, Nilsen H, Pirinen E, Andressoo JO. Nicotinamide riboside first alleviates
symptoms but later downregulates dopamine metabolism in proteasome inhibition
mouse model of Parkinson’s disease. Heliyon. 2024 Jul 9;10(14):e34355. doi:
10.1016/j.heliyon.2024.e34355. PMID: 39108921; PMCID: PMC11301377.